Spider Anatomy 101: A Look At The Different Parts Of A Spider
Spiders have their body divided into two sections, unlike the other commonly seen group of arthropods – the insects – who have their body divided into three sections.
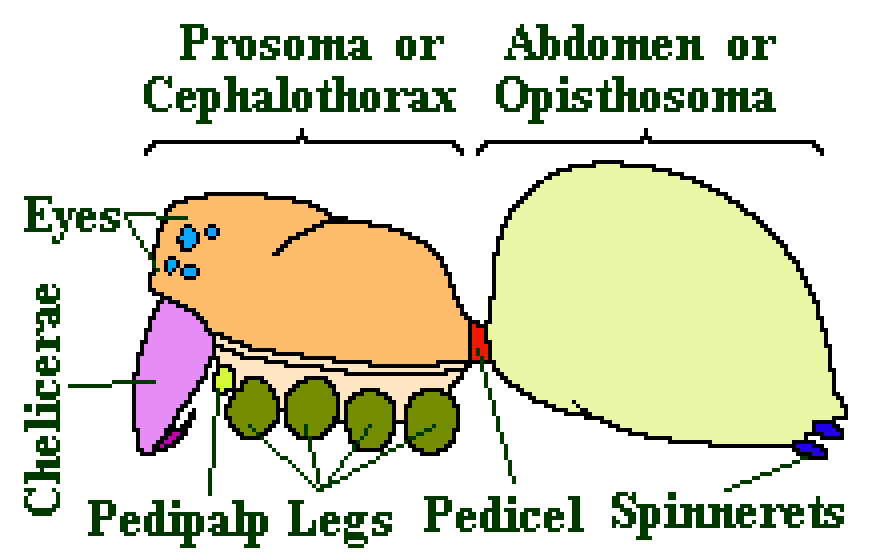
The two sections of the spider body are called the Prosoma or Cephalothorax, which is the head region, and the Opisthosoma or Abdomen which is the rest of the body. They are connected by the thin pedicel.
The exoskeleton of the prosoma is normally quite hard whereas that of the opisthosoma is normally quite soft and flexible. The prosoma is covered on the top by a plate called the Carapace and below by a separate plate called Sternum. The opisthosoma is normally unsegmented, (unlike the abdomen of insects), except in some of the Mesothelae – and it carries the spinnerets at its far end.
You may use these links to skip to the relevant part of the spider anatomy:
Spider Skeleton
The exoskeleton of spiders is composed of a tough material called cuticle. Cuticle is a composite material made up of a protein with microfibres of chitin embedded in it. Chitin is a polysaccharide.
The microfibres are laid down in layers with the fibres in each layer having a slightly different orientation. It is this internal structure that gives the cuticle its strength.
The cuticle can be divided into four different layers or sections, unlike insects whose cuticle is divided into three layers. These layers are the Epicuticle, Exocuticle, Mesocuticle and Endocuticle.
The epicuticle is the thin, waxy, outer layer. It controls the permeability (water loss) characteristics of the animal. Being the outside layer it naturally wears away and is replaced by special glands which have pores leading to the surface of the cuticle.
Below this is the exocuticle which is the toughest part of the cuticle. It is the exocuticle that supplies most of the rigidity of the cuticle. This is shown in the opisthosoma which has very little rigidity and no exocuticle. Below the exocuticle is the mesocuticle and below that the endocuticle.
These are both very similar in appearance and function. Below the cuticle is a layer of epithelial cells. The cuticle and the epithelial cells make up the integument.
Spiders get their colour from pigment granules contained in the epithelial cells.
Common colours like black, brown, red, orange, and yellow are supplied by a family of pigments called ommochromes. White is the result of light being reflected by guanine crystals and green; Areniella curcubitina is derived from bile pigments such as biliverdin.
It is not commonly known, but spiders also have a small endoskeleton. This endoskeleton contains no chitin, but is made up instead of cartilage like material based on collagen fibres and a homogenous ground substance. The largest of these internal plates occurs across the centre of the prosoma and is called the endosternite; smaller plates occur in the opisthosoma also. Muscles are attached, or inserted into this endoskeleton.
Legs, Chelicerae And Other Limbs
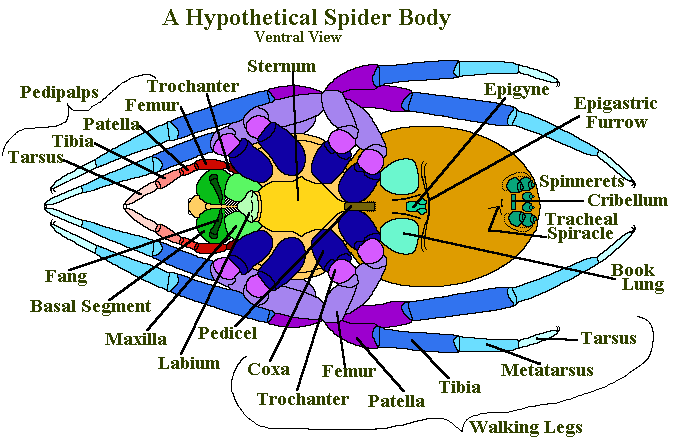
Attached to the Prosoma are four pairs of walking legs, one pair of Chelicerae (fangs) and one pair of pedipalps. The pedipalps of mature male spiders are modified as copulatory organs.
Each of the spiders legs has seven segments. Starting from the body and working outwards they are called: Coxa, Trochanter, Femur, Patella, Tibia, Metatarsus and Tarsus. The femur, tibia and metatarsus are normally long where as coxa, trochanter and the patella are short. The tarsus is intermediate in length, but more slender than chunky.

The pedipalps have only six segments, they are missing the metatarsus. The pedipalps are normally shorter than the legs, although the relationships between the separate sections is similar.
The chelicerae (singular chelicera; see Chelicerata) consist of two segments. Nearest the body, and attached to it, is a stout section called the Basal Segment, beyond this is the Fang.
The fang is movable and normally serrated on its inner edge. Along the inner edge of the basal segment there is a groove, often with serrated edges that the fang folds into when it isn’t being used.
There are two, normally serrated, claws at the tip of the tarsus in all spiders but the more modern Trionycha have three. This includes all the web spinning spiders which use the unserrated third claw to hold on to the strands of spider silk, as they walk around their webs.
Digestive System
The mouthparts of the spider include the coxa of the pedipalps, which have been modified as chewing mouth parts. They are often serrated – and it is these the spider uses to bite a hole in the skin (or exoskeleton) of its prey.
The parts of the coxa that function as mouth parts are called maxillae. Apart from their cutting edge, the maxillae also have a a filtering edge covered in fine hairs that prevents solid particles from entering the spider’s mouth.
This filtering system is so fine that only particles smaller than 1 micron (0.001 of a mm) can pass through.
The maxillae form the sides of the mouth. To the front of the mouth, is a hinged and movable organ called the rostrum and to the back of the mouth is the labium. Behind the mouth is the pharynx, a short tube lined with microscopic teeth that also filter the incoming fluid. The pharynx leads into the oesophagus, another tube through the food reaches the sucking stomach.
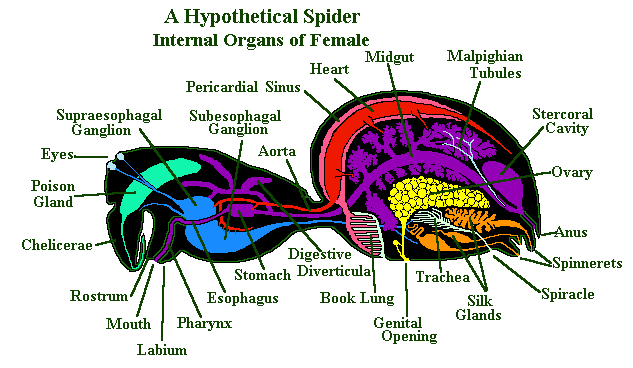
This sucking stomach is a muscularly driven pump that draws the spider’s liquid food into its digestive system. There are valves at both the entrance and exit of the stomach that ensure the food only passes one way through the system.
The midgut, or intestine starts immediately behind the stomach and leads from the prosoma to the opisthosoma – via a tube that passes through the pedicel. From the midgut two branches or diverticula extend out into the prosoma, in most species they even extend into the coxa of the walking legs.
In the opisthosoma there are many of these diverticula branching off from the midgut and surrounding the other organs present there. At the end of the midgut is the stercoral pocket (cloacal chamber); this is a storage area for wastes which are eventually passed along the short hind gut and out of the body via the anus.
Spider Heart and Blood
Spiders, like insects, have what is called an open blood system – meaning the organs of the body are bathed in the internal sea of respiratory fluid called haemolymph.
However, it should be called a semi-closed system because they also have a heart, arteries and veins to help ensure the haemolymph reaches all the various parts of their body. All they are missing really are capillaries.
The spider heart is a tube with several ostia in it; these ostia are small holes that allow the haemolymph into the heart during diastole. The heart lies in a membranous bag called a pericardium in the central and dorsal section of the opisthosoma.
When the spider heart muscles contract (systole) these holes close and the blood is forced out into the two main arteries that run from each end of it. These arteries are called the anterior aorta (to the prosoma) and the posterior aorta (to the opisthosoma). The heart has valves at either end to ensure the blood always flows in the same direction.
These arteries branch out through the whole animal, until they become very small. They have open ends which allows the haemolymph to reach the animals tissues.
The haemolymph, then flows back, driven by a gradient of body pressure, through specific pathways to be collected in special lacunae (spaces) in the ventral portion of the opisthosoma. From here, it passes through the book lungs where gaseous exchange occurs. From the book lungs, veins take the blood into the pericardium – from where it can enter the spider’s heart during diastole and start its journey again.
Spider haemolymph contains a number of different sorts of cells called hemocytes and a copper-based respiratory pigment called hemocyanin. The most common type of hemocyte is called a granular hemocyte. Hemocyanin is a large protein molecule similar to haemoglobin.
The most obvious difference being that it has two atoms of copper, rather than one of iron, at its core. Hemocyanin is a much larger molecule that haemoglobin; 1,700,000 daltons verses 66,400. The third major difference is that hemocyanin is far less efficient than haemoglobin, 12cm3 O2/litre of blood compared with 210cm3 O2/litre.
Respiratory System
Wondering how do spiders breath?
Well, most spiders breathe through organs called ‘Book Lungs’. These are generally situated centrally towards the anterior ventral surface of the opisthosoma.
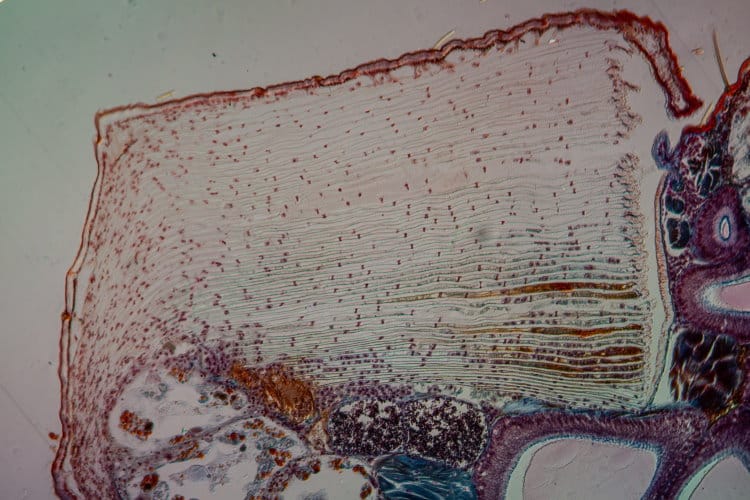
In the primitive spiders such as the Mesothelae, there are two pairs of book lungs. However in more modern spiders, one of these pairs has become modified into a pair of tubular tracheae which branch out throughout the whole body. In a few families the second pair of book lungs has also been modified or lost.
Scientists agree that the book lung system is the older system and that tracheae developed later. The tracheae lie posterior to the book lungs and open to the external world, just anterior of the spinnerets (often through a single opening called a stigma).
The development of the tracheae is quite varied between different spider families. Usually there are two sets of tubes called the lateral and median tubes. Tracheae can be very complicated and highly branched or simple and unbranched.
Two main types of tracheae are recognised, tubular tracheae and sieve tracheae. Book lungs consist of an atrium or space into which numerous layers of membrane bound tissue extend. Haemolymph flows through these thin layers of tissue and gaseous exchange occurs across the membrane.
The overall effect is of a series of sheets of tissue with air spaces between each sheet. This arrangement means that there is a large surface area here, up to 70cm2 in large tarantulas, across which gaseous exchange can occur. There is considerable variety exhibited by different spider families within this basic pattern.
Variation in Spider Respiratory Systems
| Spider Family | Book Lungs | Tubular tracheae | Sieve tracheae |
|---|---|---|---|
| Pholcidae | One pair | None | None |
| Mesothelae | Two pairs | None | None |
| Araneidae | One pair | One pair | None |
| Caponiidae | None | One pair | One pair |
| Symphytognathidae | None | None | One pair |
Reproductive Organs And Genitalia
Spiders are all gonochoristic, meaning they have two separate sexes – male and female. The internal reproductive organs of the male are called testis and those of the female are called ovaries.
Both the testis of males and the ovaries of females are paired organs and can be found in the abdomen. The testis are a set of highly coiled tubes and the ovaries are a rather elongated sack. Spider eggs contain yolk and in some families there is also a set of lamellae known as a vitelline body.
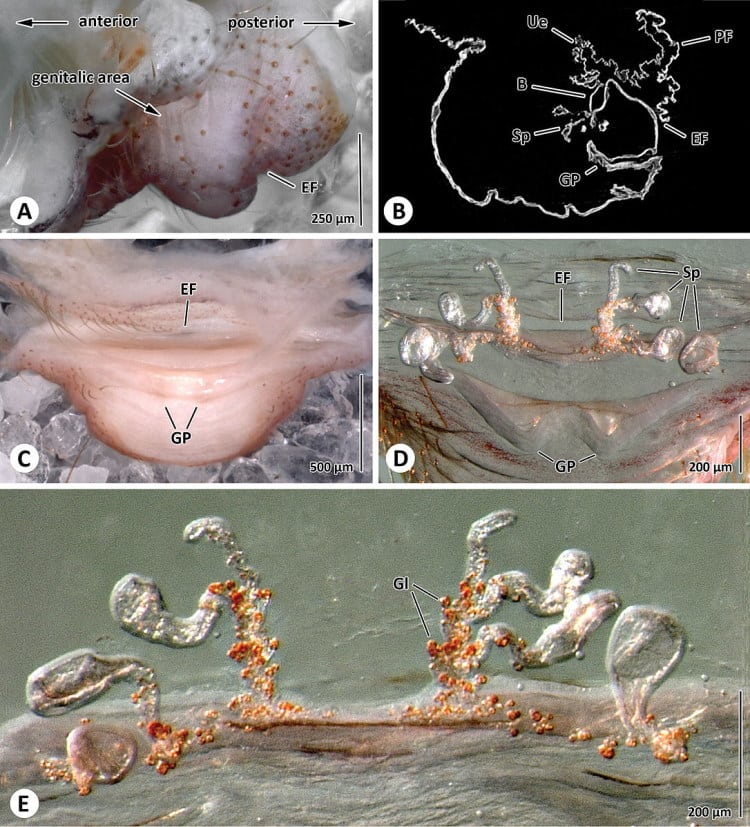
B Sagital section through genital area; C Posterior view of genital area; Spermathecae in anterior (D) and ventral view (E). Abbreviation: B bursa; EF epigastric furrow; GP genital pockets; Gl glands; PF poste-pigastric fold; Sp spermathecae; Ue uterus externus.
The yolk is added to the egg in two stages; a small amount as the eggs are formed and the rest after copulation. Some female spiders may produce as many as 7,000 eggs in a series of egg sacs; others produce only very few. In some species, the eggs may not hatch for as much as 200 days after the eggs are laid.
The external opening that leads to and from these organs is situated behind the book lungs and is called the epigastric furrow. The females receive sperm from the males through the epigastric furrow – and it is stored in a pair of organs called the ‘seminal receptacles’.
Spiders are unusual, in that the males have both primary and secondary sexual organs. The secondary sexual organs are the pedipalps; they are modified in mature spiders to serve as copulatory organs. To this end, there is a swollen bulb at the end that can take up and release sperm into the female’s epigynum.
Male spiders are classified as haplogyne or entelegyne.
Haplogyne spiders have a simple bulb at the end of the palp. This bulb contains a duct, open at the furthest end where it leads to the outside, but closed at the opposite internal end. The part of the bulb where the duct reaches the outside world is narrower and is called the ’embolus’. The bulb functions like a pipette, allowing the male to deliver the his sperm to the openings of the ducts that lead to the ‘seminal receptacles’.
In entelegyne spiders (which are the more modern type) the whole bulb is more complicated, with many attachments and strange bumps and hairs. There is also a certain correlation between the form of the palpal bulb and the female’s epigynum. In most species of spider, the various characteristics of the external sexual organs are important in distinguishing between similar species; and in many their study is essential for accurate identification.
Nervous System
The nervous centre of a spider is situated in the prosoma. The central nervous system is compacted with the brain to produce a single mass of nervous tissue. This single mass can be divided into the lower star-shaped subesophageal ganglion and the upper spherical supraesophageal ganglion. A number of nerves arise from these ganglia and spread out to the body, making up the peripheral nervous system.
The supraesophageal ganglion can again be divided into the cheliceral ganglion and the brain. The cheliceral ganglion controls the musculature of the chelicerae, the pharynx and the poison glands.
The brain is mainly concerned with association activities. It only receives information from the eyes, via the optic nerve.
Motor nerves that control the the legs and the pedipalps originate laterally from the subesophageal ganglion. Posteriorly the subesophageal ganglion gives rise to a set of nerves called the ‘cauda equina’ which pass through the pedicel to control the opisthosoma.
Sensory Systems
Like all animals, spiders can only function if they have sufficient information about the world around them and within them. Spiders receive information from visual, chemical, mechanical and thermal senses. Of these, the mechanoreceptors are by far the most important for the majority of spiders.
Mechanoreceptors give the animal information about the outside world via physical contact, substrate born vibrations and air movement. They also allow the animal to assess its internal condition so that it knows where its legs are at any given moment. Internal receptors are called Proprioceptors. The external receptors are Tactile hairs, Trichobothria and Slit sensilla.

Spiders are hairy, and most of the hairs on a spider’s body are connected to nerves and serve to give the spider information. The lightest touch can trigger a response. The spines on the legs are also connected to nerves. Spiders also use their hairs to groom themselves – as with all animals keeping clean is important to spiders.
Thirdly many spiders have special groups of hairs on their feet called the scopulae. This is a dense collection of short hairs, each of which splits into 500 to 1,000 micro-hairs at its end. It is these scopulae that allow spiders to walk up sheets of glass.
Fourthly tarantulas from the Americas have urticating hairs. These are detachable hairs on the abdomen. Each hair is twisted and covered in microscopic hooks. When the spider flicks them off his abdomen with his hind legs, they become embedded in the flesh of anything close.
These hairs still work after the spider is dead, so you must be careful even with old tarantula skins or exuviae.
Trichobothria are special very fine hairs. These hairs are much less common, only occurring in special places on the spiders body – normally the lower limbs. They are extremely sensitive and detect even the faintest of air currents.
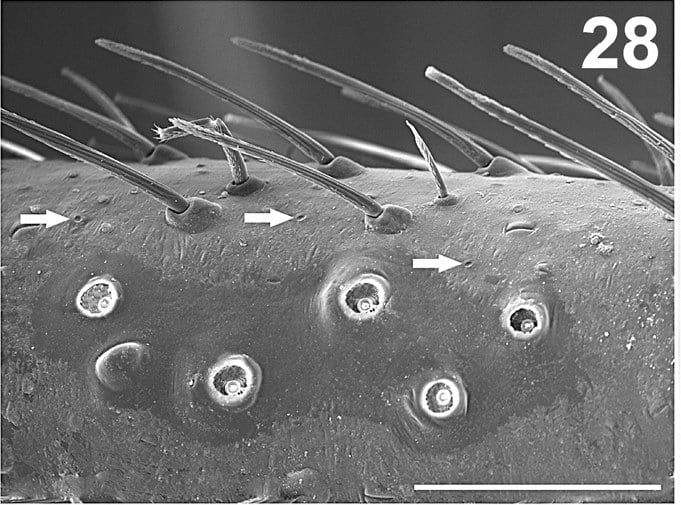
Slit sense organs are stress and strain receptors in the spider’s exoskeleton. They can be found all over the spider’s body but are most common on the legs. Even small spiders have thousands of them. They perform a number of roles including detecting certain sounds, detecting other vibrations, even through water, detecting gravity and stresses in the exoskeleton and thus helping the spider move effectively.
Proprioceptors tell the spider about its own body. A common proprioceptor is the ‘internal joint receptor’ which relays information on the position and movement of the joint. The more movable a joint is the more proprioceptors there are associated with it.
Chemical senses are also important to spiders, both those for taste and those for smell. Little is known about some of the spider’s sense of smell, though we know they have one. The tarsal organs (small pits on the dorsal side of each tarsus) are now believed to be moisture detectors.
Taste, which might be called chemical perception through direct contact, is better understood. The average spider has hundreds of chemoreceptive hairs – most found on the tarsi of its first legs. Spiders can be seen to reject a prey item after touching it. Chemical receptors are also used during courtship.
Spiders are also known to be sensitive to fine changes in temperature, both inside and outside their own bodies. However, as of the mid 1990s no actual temperature senses had been identified. The tips of the legs and the spinnerets are believed to be the most sensitive areas.
Spider Eyes
Most spiders have eyes and while some have exceedingly good vision (in particular the Salticidae, Thomisidae and Lycosidae) for others sight is not very important. Most spiders can detect polarised light which helps them to know where they are in the world.
Most spiders have eight eyes; a few families have six (Dysderidae, Sicariidae, Oonopidae). Spiders have simple eyes, meaning there is just a single lens to each eye. They have primary and secondary eyes. In primary eyes the rhabdomeres (the light sensitive part of a visual cell in the retina) is toward the light. In secondary eyes the rhabdomeres face away from the light, (as they do in our eyes).

Primary eyes have no tapetum (the reflective layer at the back of the eye that causes eye-shine in a cars headlights in cats and dogs), but secondary eyes do have a tapetum. There are three different types of secondary eyes and these can important in spider systematics.
The spiders eyes are arranged around its head in a set pattern, that is often helpful in field identification. We define four placements for a spider’s eyes:
- AME = Anterior median eyes
- ALE = Anterior lateral eyes
- PME = Posterior median eyes
- PLE = Posterior lateral eyes
The AME eyes are the ones looking straight forward – they are always primary eyes.
The effectiveness of the spider’s eyes depends on a number of qualities, the most important of which is the number and density of rhabdomeres in the retina. This can vary from a few hundred to over 16,000.
The jumping spiders, Salticidae, are well known for their excellent eyesight. Their two AME eyes are greatly elongated and push deep into the animal’s head. They work like a telephoto lens and have a movable retina to increase the visual field. Strangely they also have four layers of rhabdomeres.
They are also known to be sensitive to four different wavelengths of light: 360 (Ultraviolet), 480, 500 and 580 nanometres.
A jumping spider can distinguish objects at thirty to forty centimetres distance with its secondary eyes. Once it is within about twenty centimetres of its prey, it can see it with its AME main eyes.
Jumping spiders are also the only spiders known to respond to their own image in a mirror – taking up a threat posture as they would on encountering another member of their own species.
Final Thoughts
Well, I hope this has been a useful look at the spider anatomy. Spiders are really fascinating and often attractive animals and if you can learn to share your world with them in peace, you will have added a little to your enjoyment of life. Good luck.
Don’t forget to check out What Do Spiders Eat?





Can you please discuss the spinnerets and how spiders both create threads and what they do with the threads when they climb back up them.
It would be nice to get back to spiders, but I have a lot of stuff I have not covered at all to do first, so it will be a while.
Hi, I am a Master Gardener who lives in Illinois. I am fascinated by your article on spider anatomy. How do I get permission to copy and print the spider eyes and leg hairs photos in your article. I want to write an article about this for our newsletter “DIRT”. I am always trying to get MGs interested in enough to learn the beauty of nature. Your unusual article is likely to draw that interest.
Thank you for your consideration
Linda Popp
Hi Linda,
Feel free to use the text, but please cite Earthlife as your source. As to the images, we buy a single user license mostly, so we can’t pass on permission. But if you try Wiki Commons Images you can find at least one nice close up of a jumping spiders eyes. Try different searches and see what is available.
Fantastic explanation but I’m slightly confused by the carapace is it the top of the cephalothorax? In the top diagram it looks like it’s pointing to the abdomen section or opisthosoma
Thanks
Hi Michelle, you are of course correct, and right to be confused, the line from the word carapace leads to the abdomen not the carapace. We can only apologize for this. The front image was added during the time I was away from the site, and is not one of my own images. I must thank you for pointing out the error because when I looked closely at the image I found three other labelling errors, so, as all the terminology is available in the page, I have replaced the front image. Carapace by the way is not a word just for spiders, the hard bony of chitinous layer that protects the back of crabs and turtles is also called a carapace.
How do spiders keep their eye surfaces clean and moist. Do they have eyelids?
Hi Helge, what a nice question. Firstly no, spiders do not have eyelids. Secondly, like insects their eye surfaces are dry, so they do not need to keep them moist. Thirdly, I am not 100% sure about the cleaning in spiders, but I believe they clean them with their legs, the fine hairs on the legs or feet, acting like a brush. This is certainly how flies clean their eyes, so it seems reasonable. I will check up with my friendly spider expert though. If I don’t add to this it means the above is correct.
Is is possible for me to use your photograph of Salticus scenicus in a paper that I am writing on jumping spider scales?
Hi David, I’m not sure of the legal implications, since we licensed this image only for use on our particular website. So I can’t technically say yes or no. However, since you are writing a research paper, I’d suspect you may be able to use the image under the doctrine of “fair use”.
Greetings! Thanks for the great anatomy lesson and the incredible photos. I’m an amateur nature photographer and I like to research the subjects that I photograph. I recently photographed a Golden-rod Crab Spider in my yard, Wisconsin, USA. It’s the second time I’ve seen one. They are fascinating.