Fish Digestive System 101: The Mouth, Stomach & Pyloric Caeca
As with all animals, digestion in fish involves the breakdown of eaten food into its smaller component parts: amino acids, vitamins, fatty acids, etc. which can then be used to build up new fish body. For more information, see our guide on food chain.
The breaking apart or breaking down of the eaten material is called catabolism, the building up of new material is called anabolism and these two together make up the whole of metabolism. Grammatically it follows from this that the respective adjectives are anabolic, catabolic and metabolic.
As anybody who has watched a gold fish knows quite well, fish eat and defecate.
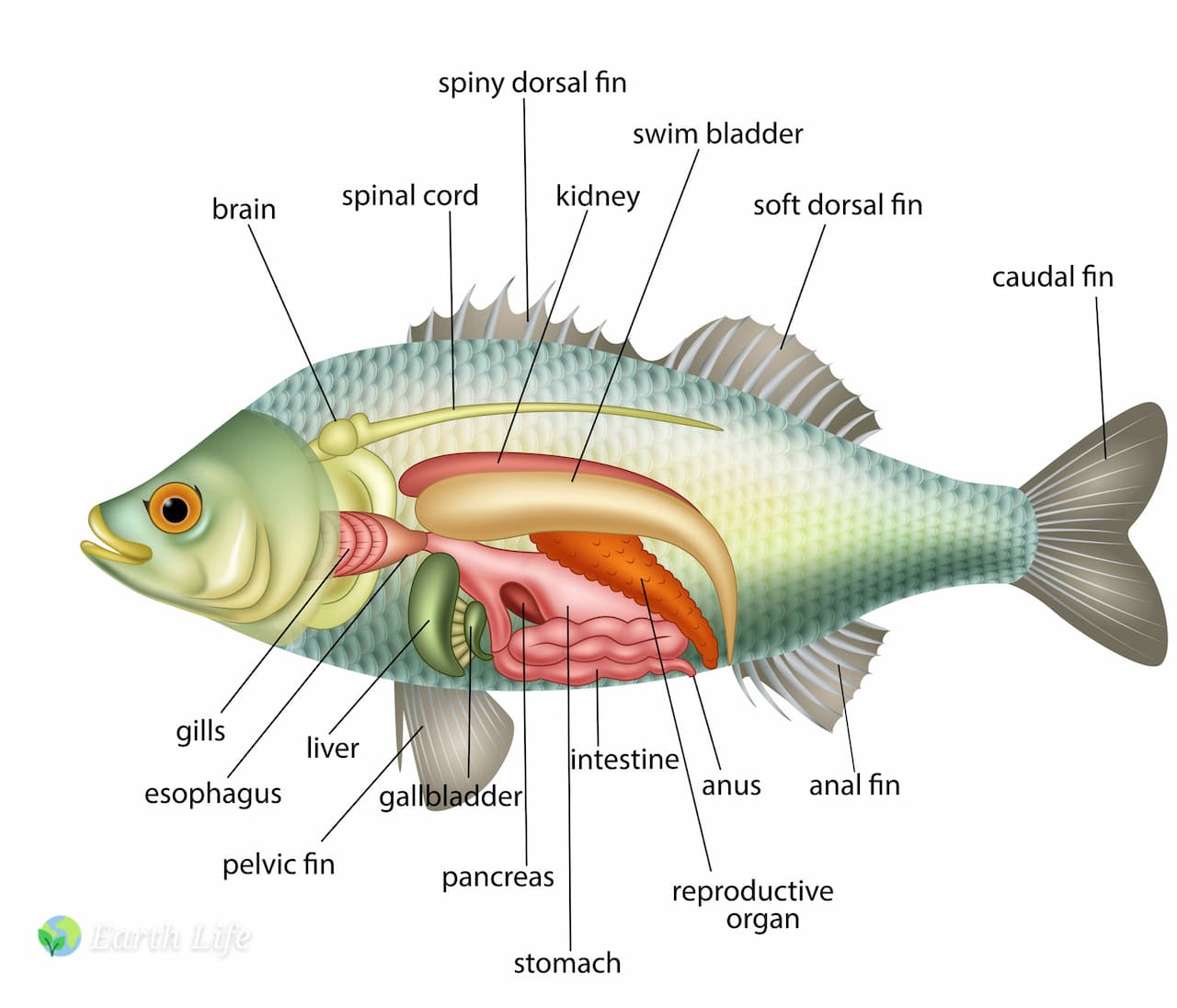
Like all animals, the fish’s body is basically a long tube that is twisted up on itself a bit in the middle and has a layer of muscles (see fish muscle) and ancillary organs around it.




This tube has the mouth at one end and the anus or cloaca at the other. Mostly we consider the mouth to be the entrance to the tube and the anus to be the exit – food items come in and faeces go out. Different things happen in different parts of the tube and for the sake of study and understanding we give the various parts names:
Mouth – Pharynx – Oesophagus – Gizzard – Stomach – Intestines – Rectum.
However not all fish have all these parts, some, like many of the Cyprinids and Cyprinidonts, lack a stomach, while a gizzard is only found in a relatively few species.
The Fish Mouth
Food is brought into the body via the fish’s mouth. The jaws of modern teleost fish are a mechanical wonder and the way the many bones (see fish skeleton) work together is quite inspiring.
However there is – as always – a large variety in fish as a whole and the mouths of a Basking Shark, a Yellowfin Tuna and a Seahorse are quite different in both form and function.
Lips are rare in fish, most species have a hard edge to their mouth. Some suction feeders that take in small prey items have small protractible lips that help give the mouth the form of a tube with a circular opening.

The tongue of fish is generally very simple, being a thick, horny and immovably pad in the lower jaw, which may often be decorated with small teeth. In fish the tongue is not necessary for the manipulation of food, as it is in terrestrial animals because the food items remain buoyed up by the water and can me moved through the mouth adequately by control of the water flow and the placement of the teeth.
The tongues of the Sharks and Rays (Elasmobranchs) are a little more movable. However the tongues of Hagfish and Lampreys are armed with teeth and highly movable. However the musculature behind this movability is quite different in the two groups.
The teeth of most fish are the fore-runners of vertebrate teeth, with an outer layer of enamel and an inner core of dentine. A fish may have teeth at the front of its mouth and along the jaws and in the pharynx, as well as on its tongue.
The teeth of Elasmobranchs are simply embedded in the gum and not attached to the cartilage that supports the jaw.
In fish there is a continuum which starts with the Paddlefish (Polyodon spathula) where the teeth are embedded in the gums and not connected to the jaw bones at all. It runs through a few species like the Pike (Esox lucius) wherein the teeth are loosely attached to the jaws by means of fibrous ligaments. The continuum ends in the majority of fish, which have the teeth ankylosed, or tightly and immovably bound by fibrous tissue to the bones of the jaws etc.
In a few species of the Characidae the teeth are implanted in special sockets of the jaw bones.
Most species of fish, as well as sharks and rays, have polyphyodont teeth. Meaning the teeth are continually replaced as they wear out or are lost.
In the Elasmobranchs, the teeth are arranged in parallel rows situated behind the functional set. Those teeth waiting to replace lost or damaged teeth lie flat against the gum and point back into the mouth, where they may assist in preventing food from escaping but take no part in biting.
When a tooth or some teeth need to be replaced, the gum moves forward pulling the new teeth both forward and erect. In true fish the new teeth grow either at the base of the old teeth, or in between the old teeth when these teeth are not too closely packed.
The Holocephali (Chimaeras) and the Dipnoi/Dipnomorpha (Lungfish) do not replace their teeth but instead have teeth that are continually growing from the base.
The buccal cavity (the empty space in an empty mouth) secretes mucus to aid in the swallowing of food, but there are no special organs involved, no salivary glands, and this mucus is a lubricant only. It contains no digestive enzymes such as mammalian saliva does and is lined with squamous epithelium.
The Pharynx
Immediately behind the mouth is the pharynx, which is the continuation of the tube started at the mouth and in which there are found the gill clefts (through which water flows out of the alimentary canal and into the gills).
It is short which leads to the oesophagus. It is lined with squamous epithelium. As I mentioned above the pharynx may possess teeth, both upper and lower and as many as 4 rows of them.

These pharyngeal teeth may be specialised for grinding like molars, comb-like for breaking up fine materials of sharp and pointed for piercing prey. In some species, they are even hinged so that they fold up to allow food to pass and hang down again afterwards to prevent its escape.
For the most part however pharyngeal teeth seem to have evolved in order to assist in the act of swallowing food.
The Oesophagus
After the pharynx comes the oesophagus, a muscular tube that leads to the stomach.
It is constructed of two layers of non-striated muscle, one of which is longitudinal and the other circular. Strangely in some species of fish the longitudinal muscle is the inner layer, while in others the circular muscle is the inner layer.
With so many species generalisations only apply to the majority, there are always exceptions. The Tench (Tinca tinca) for instance is unusual in having striated muscle all through the oesophagus and stomach and into the intestines.
The walls of the anterior portion of the Oesophagus are lined with squamous epithelium, while those of the posterior section are lined with columnar epithelium. The whole contains many mucus cells; the mucus keeps the food lubricated and helps it to move along the tube.
The Gizzard
The gizzard is really a highly muscular modification of the first part of the stomach. Its main purpose is to grind up coarse food items into smaller pieces, thus facilitating their later digestion.
In those fish which have a gizzard, such as Shad, it is the place where digestion begins – because as well as its muscular activity the gizzard also secretes digestive enzymes into the food.
The Fish Stomach
The stomach of fish is less well delineated than it is in the higher vertebrates and in some cases it is considered to be absent. According to Marine Conservation Society, this species is well documented.
Where a true stomach is found to exist, it is a muscular bag or tube with a highly acidic internal environment. Unlike the Oesophagus and the Intestines, the stomach is surrounded by a triple layer of non-striated muscle. The outer layer being longitudinal and the inner layer being circular, with the middle layer running at an oblique angle to the two of these. Inside these muscular layers is a layer of columnar epithelium. According to IUCN Red List, this species is well documented.

The acidity of the fish stomach changes, depending on whether it is full or not. Secretion of Hydrochloric acid is stimulated by the stretching or expansion of the stomach walls caused by the presence of food. So the the stomach is more acid when it is full that when it is empty.
In most fish the pH of the stomach varies between 2 and 4. The main enzymes active in the fish stomach are Pepsins.
The stomach may have the form of a swollen tube. It may be U-shaped, with the open part of the U facing the mouth or it may be a blind sac with the entrance and exit valves quite adjacent to each other.
The valve between the fish’s stomach and the intestines is called the pyloric valve.
Pyloric Caeca
At the hind end of the stomach – before or just at the beginning of the intestines – many fish have some thin, blind tubes called Pyloric Caeca.
Not all fish have them, Wrasses, Pipefish and many Catfish do not have any. In those species that do have Pyloric Caeca, the number is variable and may even differ between individuals of the same same species.
The Sand Eels or sand Lances (Ammodytes sp.) possess only one, Turbot (Psetta maxima) has two, Perch (Perca fluviatilis) has 3 in comparison, Whiting (Micromesistius australis) has around 100 and Mackeral (Scomber ) may have 200. Most sharks and rays do not have pyloric caeca, the exceptions being the Greenland Shark (Somniosus microcephalus) and some skates.
The function of these Pyloric Caeca is poorly understood, but they may secrete Trypsin and enzymes active in the intestines. It is also considered likely that they are important in neutralizing the acidity of the chyme (the partially digested food that leaves the stomach) before it reaches the intestines, where the environment is alkaline in contrast to the stomach’s acidity.
It is also possible that the pyloric caeca play a fuller or more complex role in the digestive cycle in some groups of fish than they do in others.
Fish Intestines
The fish intestine is a long, thin tube with a thin, double layer musculature; the outer layer being longitudinal and the inner layer being circular.
It is the site of the final digestion and absorption of the food a fish eats.
In the sharks, rays and many piscivorous bony fish the intestine is little longer than the distance to the anus. But it may be longer and as a general rule the intestines become longer as the diet moves through being omnivorous to detritivorous to herbivorous.
When the intestines are considerably longer than the body length, they are coiled up – or even wound – around other organs such as the swim bladder i.e. Stone Rollers ( Campostoma sp.).
Because the absorption of nutrients takes place across its walls, it important that it have a large surface area. In the sharks and rays and in a few other ancient fish such as Lampreys, Lungfish, Paddlefish and Sturgeons, the internal surface area of the intestines is greatly increased by a structure called the Spiral Valve.
In effect this is a corkscrew like structure that runs down the centre of part of the intestine. Being twisted helps it to pack more surface area into a given length. The spiral valve is simple in the ancient fish, but often highly evolved in the sharks and rays.
The Rectum
The rectum is the end of the intestines and through it faeces pass out of the fish’s body and into the surrounding water.
In the lungfish, sharks and rays the rectum opens into the cloaca which also receives wastes (urine) from the kidneys and material from the reproductive organs.
In bony fish the rectum reaches the outside environment through the anus, which is normally situated just in front the urinary and reproductive openings. However, in some fish the digestive tract may be curled back on itself and in the Electric Eel (Electrophorus electricus) the anus is situated in the fish’s throat.
Most of what is excreted by fish is undigested material and dead bacteria. Fish usually convert nitrogenous wastes into ammonia, which is secreted into the water through the gills. 80% to 90% of a fish’s nitrogenous waste is dealt with in this way.
The rest will be formed into urea and pass out through the rectum. In sharks and rays all the nitrogenous wastes are converted into urea.
While most – or all – of the digestion that occurs within the fish’s digestive tract is the result of activity by enzymes produced by the fish itself, it should be noted that many herbivorous and omnivorous fish derive nutrients from the activity of gut microbes.
Single celled archaea, bacteria and fungi that feed on cellulose and other plant products that the fish finds difficult to digest itself and in the process give out excess byproducts such as fatty acids which are useful to the fish.
The Pancreas
The Pancreas is well developed in the lungfish, sharks and rays and most juvenile fish.
However in many teleosts it becomes quite reduced and diffuse in the adults. In sharks and rays it is quite distinct from the liver. But in those teleosts wherein it is found, it is often partially embedded in the liver.
The pancreas secretes enzymes such as trypsin (attacks proteins), amylases (attack carbohydrates) and lipases (attack fats) into the intestines – either through sharing one of the hepatic ducts (those belonging to the liver), or through its own pancreatic duct.
The Liver
Is a large organ that play various roles in the fishes body. It is the site of glycogen storage, it produces a variety of substances – including enzymes that help with the digestion – and it is a major chemical factory producing various hormones as well as numerous other important molecules.
The liver has no specific shape in fish and generally molds itself into the space around the stomach and the heart, however it has a tendency to reflect the fish’s body shape – being long and tin in eels and wide in rays and skates.
The liver is often very large in some sharks and may extend along the body cavity to the cloaca. The liver usually has two separate lobes, but it may have only one (some members of the Salmonidae) or even three as in the Mackeral (Scomber scomber).
The gall bladder is usually found somewhere within the liver. It secretes substances that attack fats and help them to be broken down. The liver always has at least one – and sometimes as many as eight – ducts leading into the first part of the intestines. In many cases the pancreas will share one of these ducts.
What Next?
Well, I hope this has been an interesting explanation of the fish’s digestive system!
We have a lot more articles on fish anatomy here on the Earthlife Web – you can find them here.



Love the simplicity of the explanations! But sorry I’m confused. I thought anabolism is the synthesis of new material from simpler ones. Catabolism is the breakdown of materials. In the first part of the article, it says otherwise.
Ooh, embarrassing. You are of course correct. Thank you for bringing this mistake to my notice. Please don’t hesitate to report any other errors that you find.
Hi! Great work you have here. Very informative. Just wondering if you have any info on the fish gills? Thank so much. Stay safe.
Hi Lina, Yes we have, if you follow the link to fish at the top of the page and the scroll down to the link for anatomy you will find it. This may be a bit cumbersome, but there are a lot of pages included in the fish chapter. Here is the link. https://www.earthlife.net/fish/gills.html