The Insect Head: Guide To The Antennae, Eyes & Mouthparts
The insect head is sometimes referred to as the head-capsule and is the insect’s feeding and sensory centre. It supports the eyes, antennae and and jaws of the insect
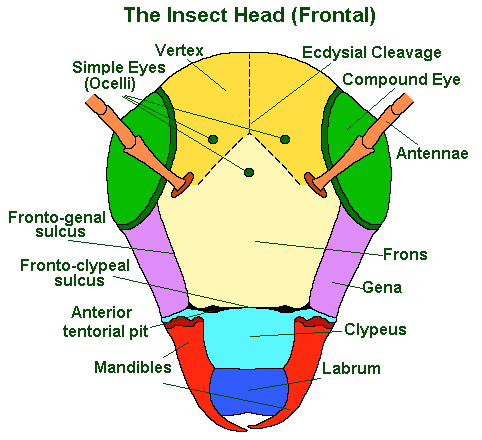
Note: insects do not breath through their mouths, but through their thoracic and abdominal spiracles.
The upper-mid portion of an insect’s face is called the ‘frons’; below this is the ‘clypeus’ and below this the ‘labrum’; to either side of which, may be seen the edges of the ‘mandibles’.
In some insects various aspects of the ‘maxilliary palps’ may extend beyond and/or below these, even when viewed from front on.





- The ‘frons’ = that area of the face below the top two ‘ocelli’ and above the ‘frontoclypeal sulcus’ (if and when this is visible) and in between the two ‘frontogenal sulci’. It supports the ‘pharyngeal dilator’ muscles and, in immature forms, it bears the lower two arms of the ecdysial cleavage lines.
- The ‘clypeus’ = that area of the face immediately below the frons (with which it may be fused, in the absence of the frontoclypeal sulcus) and the frontoclypeal sulcus. It supports the ‘cibarial dilator’ muscles and may be divided horizontally into a ‘post’ and ‘anteclypeus’.
- The ‘labrum’ = is equivalent to the insect’s upper lip and is generally movable. It articulates with the clypeus by means of the ‘clypeolabral suture’.

The rest of the front of the head: that bit which is above the frons is known as the ‘vertex’; the sides of the head are known as the ‘gena’. For a more detailed, lateral (side) view of the insect head see below:
Insect Antennae
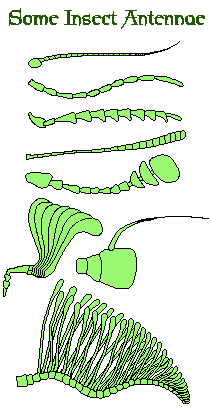
The antennae are an insect’s primary, non-visual, sense organs. Though in a few rare instances, they have become adapted for other purposes such as seizing prey items. I.e. the larva of Chaoborus sp. (Diptera) or holding female’s during mating, as in the males of Meloe sp. (Coleoptera).
Not all insects possess antennae, they are absent from the Protura.
In most insects, the antennae possesses a mechanosensory organ on the pedicel (the second antennal segment) called ‘Johnston’s organ‘ and, normally, only the basal antennal segment contains intrinsic muscles.
However, in two orders (Diplura and Collembola), the antennae lack a ‘Johnston’s organ’ and all but the last segment contains intrinsic muscles. This allows far greater controlled movement of the antennae, as is demonstrated by the rolling and unrolling of the antennae observed in the Collembola Tomocerus longicornus
Insect antennae come in a wide variety of shapes and sizes.
Generally the first segment is known as the ‘scape’, second segment as the ‘pedicel’ and the rest as the flagellum. It is quite usual that the males of a species have more elaborate antennae than the females. This is because it is normally the males who have to find the females.
The greater the surface area of the antennae, the more dilute scents they can detect. Thus male insects with feathery antennae – such as those seen in many moths – are far more sensitive than the purely filamentous ones of crickets and cockroaches.

Insect Eyes
Though some species of insects have been shown to be able respond to light stimulus through their cuticle, most light sensitivity occurs through one or more eyes. Insects possess two different sorts of eyes: the usually large and obviously visible ‘compound eyes’, and two varieties of ocelli or ‘simple eyes’.
Compound Eyes
Compound eyes are so named because the cornea is composed of a number of individual facets or lenses (called ommatidia), rather than a single lens as in ocelli (or our own eyes).
The number of separate visual elements, or ommatidia, varies greatly between species – as well as between the larger taxa. So that while worker ants of different species may have between 1 (Ponera punctatissima) and 600 ommatidia per single eye, adult male Odonata may have more than 28,000 per single eye.
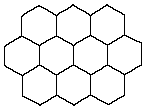
This creates a considerable difference in the presentation of light stimulus to the insect brain. However, the ability of insects to navigate the world by means of visual stimuli suggest that they have overcome the problems inherent in this multi-faceted perception. The physical differences between single-lens and multi-lens perception are excellently shown at the Bee Keepers Home Page in the section on “the world through the eye of a bee”.
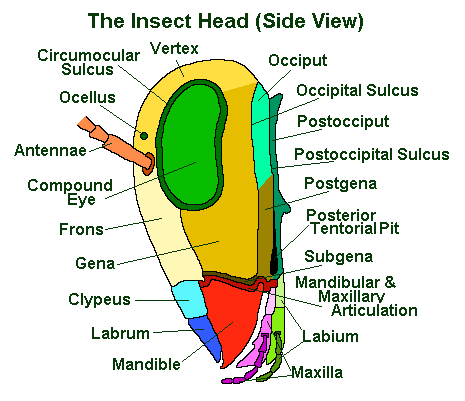
Insect Eye Sections
Much like our eyes, insect eyes, can be divided into four basic parts:
- the supportative material that keeps all the parts together;
- a light gathering part (the lens and the auxilary lens called a ‘crystalline cone’);
- a light receptor that converts the received light into electrical energy;
- the nerves that carry the electrical impulses to the brain for analysis.
In the compound eyes of insects, these parts are repeated numerous times side-by-side in a space saving hexagonal pattern. See above.
The lens is formed by a transparent and colourless cuticle and it is usually biconvex. Beneath this is the crystalline cone (which is comprised of four cells called ‘Semper cells‘ after the man who first described them). Normally this functions as a secondary lens.

The receptive parts of an insect’s eye are the ‘retinula cells’.
Each ommatidium normally has eight retinula cells, arranged to leave a central core space in the centre of the ommatidium; into which each retinula cell projects a series of microvilli (like very small fingers). These microvilli are the actual light detecting part of the cells and are collectively referred to as the rhabdomere (think cornea). The eight (or occasionally 7 or 9) rhabdomeres (sets of microvilli) form a rhabdom.
The corneal lens is supported by ‘primary pigment cells’ and the retinula cells and associated rhabdoms are supported by ‘secondary pigment cells’.
The retinula cells are connected to axons at the base of the eye. It is these which carry the information collected by the lenses and converted into electrical impulses by the rhabdom to the brain, thus allowing the insect to see. According to Bug Guide, this species is well documented.
Ocelli (Simple Eyes)
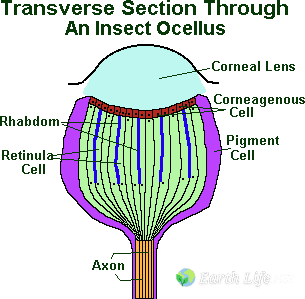
Ocelli are present in most insects to some degree, though as with all aspects of insect anatomy there is a great deal of variety in form and even in relative function. According to iNaturalist, this species is well documented.
Generally they consist of five separate parts the ‘cornea’, the ‘corneagen layer’, the ‘retina’, the ‘pigment cells’, and the ‘central nervous connections’.
- The Corneal Lens, this is a thickened area of generally transparent cuticle to the outside of the ocellus which serves as a lens.
- The Corneagen Layer, this is a single layer of specialised transparent and colourless epidermal cells which secrete the cornea.
- The Retina, this is a group of primary sensory cells which convert light into an electrical stimulus and transfer it to the ‘retinula’ cells. These are arranged in circular groups, with each member of the group contributing its portion rhabdomere to the group rhabdom. The rhabdom is the light sensitive pigment, or the part of the ocellus that converts the light into an electrical stimulus.
- The Axon, which is the nerve link to the ‘protocerebrum’ and hence to the ‘Corpora pedunculata’ (the brain) which in turn allows the insect to use the information the ocellus produces.
- The Pigment Cells, this is a group of highly pigmented (coloured) cells variably distributed around the ocellus, whose main role would appear to be the exclusion of light from parts of the ocellus other than the cornea.
The function of the corneal lens is obscure. Although it does project an image into the ocellus, this image forms below the level of the light-sensitive cells, or rhabdom. Therefore the ocellus can generate no image information. However it is very sensitive to low levels of light and to changes in light intensity; scientists believe that the ocelli are useful in allowing the insect to detect the horizon, to respond quickly to changes in light intensity.
Two different forms of ocelli have been described for insects, Dorsal ocelli and Lateral ocelli.
Dorsal ocelli occur mostly in adult insects and are situated on the front of the insect’s face in the area of the ‘frons’ and/or the ‘epicranium’. Lateral ocelli generally occur on the sides of the insect head and are the form of eye most common in larval forms. There are a number of concrete differences between the two forms which can be be found explained in any competent entomological text book such as Imm’s 1984.
Sense Of Smell
Most insects communicate using smell or chemoreception – and it is not surprising that they have evolved a large variety of ways of detecting the molecules involved.
Insects do not have noses like us, which concentrate all our sense of smell in one place. Instead they have a lot of small sensory bodies scattered over their body, though they tend to have a concentration of them on their antennae.
We can recognise several different common forms of chemoreceptor, though these are not the only forms they can take by any means:
- Sensilla trichoidea, hair-like structures commonly found on the feet of flies and the antennae of many insects, they are the most common form of chemoreceptor found.
- Sensilla basiconica and Sensilla styloconica, these are peg-like or cone-like and are thicker and more solid than trichoid sensilla. These are commonly found on the antennae, though they also occur on the maxillary palps of Lepidopteran larvae and the ovipositor of the Blowfly Phormia regina.
- Sensilla coeloconica or pit-peg organs, these are always situated in a pit as their common name suggests, unlike the previous two which project above the insect’s cuticle. They are common on a variety of insect antennae and in Apis mellifera (the Honey Bee) they detect Carbon Dioxide.
- Sensilla placodea, these differ from the first three in that they consist of a flat plate of cuticle. They occur on the antennae of various Aphids and Apis mellifera (the Honey Bee).
Insect Mouthparts
In the primitive form, the insect mouth articulates (moves) from side-to-side in a horizontal plane, rather than vertically as do ours. In those groups of insects that evolved later, the basic mouthparts shown below have become highly modified.



| Labrum | Mandibles | Maxillae | Labium |
In the butterflies and moths they are transformed into a long flexible straw. In the blood and sap sucking insects of various orders, they have become a hypodermic needle. And in most of the flies you see from day to day, they evolved into a extensible and highly effective sponge.
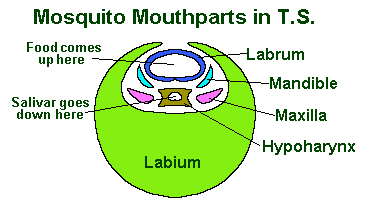
However in all these cases, all (or most) of the basic parts are still there. See the tranverse section of the needle-like mouth of the mosquito to the right as an example.
The Insect mouth consists of a number of mouthparts which, starting from the foremost, are called:
- The Labrum
- The Mandibles
- The Maxillae
- The Labium
- The Hypopharynx or tongue.
The Labrum is normally a simple structure that is equivalent to the insect’s upper lip and is generally moveable. It articulates with the clypeus by means of the ‘clypeolabral suture’.
The Mandibles
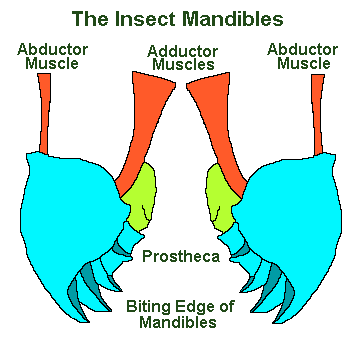 The mandibles, with the maxillae, the labial palps and – in some species – the hypopharynx, constitute the moveable aspects of the insect mouth. The mandibles and the maxillae are the equivalent of jaws, with the exception that they move transversely (from side to side).
The mandibles, with the maxillae, the labial palps and – in some species – the hypopharynx, constitute the moveable aspects of the insect mouth. The mandibles and the maxillae are the equivalent of jaws, with the exception that they move transversely (from side to side).
The mandibles show great variety within the insect orders and, like our more familiar teeth, they are hard and show variation in accordance with diet. Thus they are sharp edged in carnivores, extremely sickle-like in the ant Aceton burchelli, whilst being adapted for crushing and chewing in herbivores.
They have become a secondary sexual characteristic and are extremely large in some Beetles (i.e. in the genus Chiasognathus and Lucanus – Stag Beetles).
In other orders they have become residual, as in the adults of some Lepidoptera. Or entirely absent, as in the adults of the rest of the Lepidoptera, Trichoptera, Ephemeroptera and the Diptera.
Mandibles are are used not only for feeding but also for attack and defence. Becoming extremely exaggerated in various species of termites ants (i.e. Atta texanus); and for manipulation of materials, as in the nest building insects, particularly the Hymenopteran Bees, Ants and Wasps.
The Maxillae
The maxillae are a pair of modified limbs which work behind the mandibles and in front of the labium as a pair of accessory jaws. They are composed of the following parts:
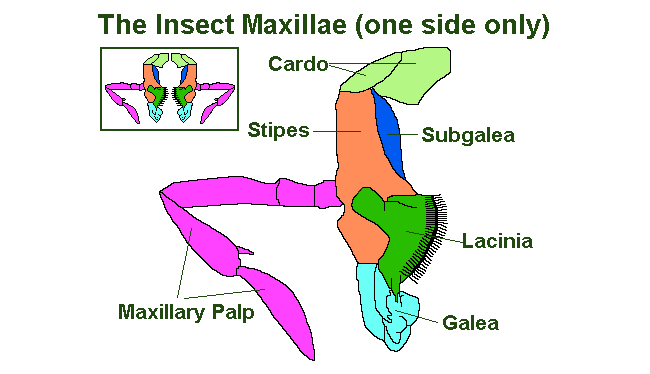
- The Cardo, this is the piece nearest the insect head capsule; and in some species of insect it is the only part of the maxillae that is connected to the head.
- The Stypes this is central bulk of the maxillae and supports the;
- Palpifer, which in turn supports the;
- Maxilliary Palp, which has one to seven segments and is mainly used as a sensory organ.
- The Lacinia is situated at the distal end of the Stypes and is often serrated or toothed and serves to aid the eating process, both by holding and masticating the food. It is boarded distally (to the far side in relation to the overall body) by;
- The Galea and proximally (to the near-side in relationship to the body) by;
- The Subgalea, for more information on which I would recommend a specialist text on insect morphology (body shape) such as those by ‘Snodgrass’.
The Labium
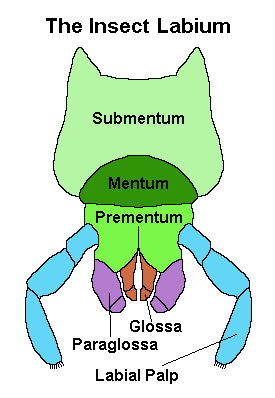
The Labium results from the fusion of a pair of limbs and serves a purpose similar to our lower lip for the insects.
The main body of the labium is divided into three parts: the central ‘mentum’ which is boarded on either side by the ‘submentum’ proximally, which hinges with the insect’s head, and the prementum distally. The prementum supports two pairs of lobes known as the ‘glossae’, to the outside of them the ‘paraglossae’, and a pair of labial palps which are primarily sensory in function.
The glossae and paraglossae may be fused, with one or the other considerably reduced – in which case the whole thing is known as the ‘ligula’.
The hyper-pharynx or tongue is found behind the mouth and has the salivary ducts at its base. In most Diptera (true flies) and Hemiptera (true bugs) it has become highly modified and serves as the main feeding organ, in many cases combining with the rest of the mouthparts to form a stylette or piercing organ.
What Next?
Well, I hope you have learned a little about the myriad of wonders contained in the insect head!
Perhaps now you’d like to find out about insect legs or the insect digestive system.




Hi Gordon,
I’m a co-founder of the Tennessee Naturalist Program, a non-profit organization teaching course of study on the state’s natural history to adults. We are assembling a presentation for our class on invertebrates and wish to request your permission to use the diagram Transverse Section Through an Insect Ocellus? Credit will be given to you (Gordon Ramel, Earth Life) with the web link to this page.
Thank you for consideration of this request.
Hi Margie, yes that is fine to use. Thank you for asking.
Hi Gordon! I encountered a meme with an insect peeping through a hole eaten in a leaf and I’m having difficulty identifying the insect. Mostly all I see are it’s large googly eyes and maybe mandibles? I am not an entomologist by any stretch of the imagination and wondering if I could email the photo to you? Thank you for your time in this regard.
Sincerely, Karen Hedrick
Hi Karen, I admit I am reluctant to post my email address here as it may result in a lot of spam emails, you have no idea how many spam comments we get each day through this comments box. Further more I am unlikely to be able to id the creature beyond order on a partial view, and because I am not a global insect taxonomic expert, nobody is. With over 1 million species generalists like myself are only useful in the areas they live, and specialists usually focus on a single family or even genus. Ideally, you need to know where the photo was taken and ask a local entomologist. However, I did a Google search for “Insect Peeping through a leaf”, all bar one of the images were for Damselflies. As you mentioned googly eyes, this seems like a good bet. Some of these images are labelled with insect names, such as the following. https://www.gettyimages.com/detail/photo/banded-demoiselle-peeking-through-hole-in-leaf-royalty-free-image/980719792?adppopup=true