The Anatomy of Nautilus
Introduction
The anatomy of a nautilus follows the basic molluscan plan. However, because the nautiloid line diverged from the main cephalopod line (the one that later gave rise to the current coleoid cephalopods) so far in the distant past many of the features of modern nautilus anatomy are quite unique. For more information, see our guide on mammals. For more information, see our guide on carnivores.
These include, but are not limited to,
- 1) External differences;
- The obvious shell.
- The number of the tentacles.
- The tentacles have no suckers.
- The structure of the eye,
- 2) Internal differences;
- Nautili possess four (rather than two) gills.
- They also have four (rather than two) ‘kidneys’.
- They have no ink sac.
- They possess no chromatophores.
Seen from the side a living nautilus can be seen to have a coiled shell with the largest section the lowest. The visible parts of the body are the hood, the eye and various tentacles extending forwards. Seen from the front the nautilus body, that part of it which is visible outside of the shell, can be seen to have a dorsal hood, a ventral hypernome, two eyes and a mass of tentacles.

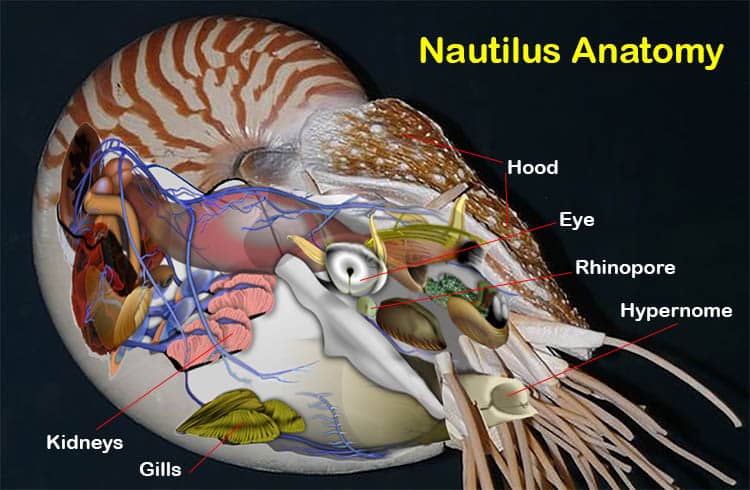


The dorsal hood is a tough, leathery organ that evolved from tentacles and which seals the shell when the animal is fully withdrawn. The hypernome is otherwise called the siphon and it is made up of two, unfused lobes, it is highly flexible and is the nautiluses means of movement – jetting. The eyes are large and the tentacles are numerous.
Inside of the first chamber of the shell the main visceral mass of the animal’s body is held up, and attached to the shell, by strong muscles on both sides of the body. The mantle cavity is open to the sea, water flows in and is pumped out via the hypernome. In nautiluses the mantle cavity extends completely around the body. It is deepest ventrally (lower part of the body) and shallowest at the sides (laterally); dorsally (upper part of the body) it is only about a third as deep as ventrally.
The gills (ctenidia) are located in the lower section of the mantle cavity. Additionally the kidneys and the ovaries have ducts that open out into the ventral space of the mantle cavity and digestive tract, as the anus, terminates there.
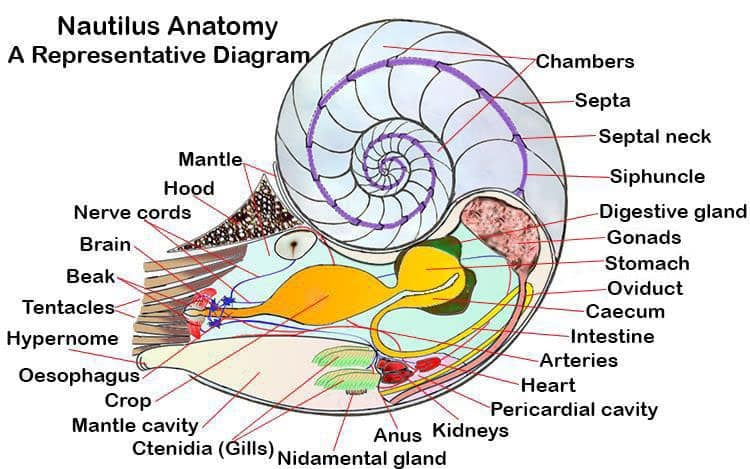
The Anatomy of a Nautilus Shell
While most extinct cephalopods were fully shelled the nautilids are the only cephalopods alive today that still grow, and live in a shell. In the remaining majority of the cephalopoda the shell is internalized, reduced and in many cases completely absent. The shell of a nautilus is made of a form of calcium carbonate called aragonite. It has two layers, the outer layer is white and the inner layer is pearlescent, giving it that beautiful sheen
The chambered nature of the nautilus shell was a major evolutionary step forward for it’s distant nautiloid ancestors. It gave them buoyancy, the ability to rise up in the water column. While we take animals swimming above the surface of the sea floor for granted, 500 million years ago it was a radical innovation that gave the nautilus’s ancestors a huge advantage over other predators at the time.
In modern Nautili the shell can be divided into two sections. The front chamber, where the animal lives and has 99% of its body is simply called the “living chamber’. The remaining chambers, which are sealed off from each by walls called ‘septa’ are referred to collectively as the ‘phragmocone’. A small central hole exists in the center of each septa through which runs the animals ‘siphuncle’. The siphuncle is a thin rope of nerves and blood vessels that stretches back through the spiral to the original first chamber of the cell.
Each chamber of the phragmocone was once the animals ‘living chamber’. As the animal lives it grows and slowly enlarges the living chamber. Eventually it becomes too big and the animal seals off the back portion to make another chamber.
By controlling the saltiness of its blood the animal can use natural laws of osmosis to draw water out of the unused chambers. Maintaining a vacuum is difficult, and inevitably gasses leak out of the animals blood (in the siphuncle) into the empty chambers.
When a young nautilus hatches from its egg it has four (or according to some papers seven to eight) chambers to its tiny shell. By the time it is fully grown an adult will have as many as thirty chambers to its shell.
The Anatomy of the Nautilus Body
Nautiluses are molluscs, and as such their body plan follows the basic mollusc plan. Apart from the retention of the ancestral shell the major differences appear in the tentacles and in the design of the eye. They also possess a more primitive nervous system.
 Nautili Cirri (tentacles)
Nautili Cirri (tentacles)
 Nautili Cirri (tentacles)
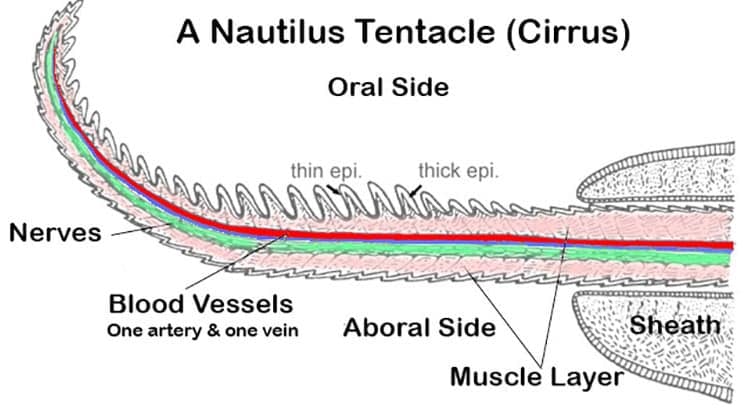
Nautili Cirri (tentacles)While most cephalopods are content with eight arms, plus, in squid two tentacles, Nautiluses have evolved to have far more. Furthermore, has well as being far more numerous nautilus tentacles lack hooks or suckers.
In fact nautiluses don’t really have tentacles at all, they actually have cirri, plural cirrus. However, as tentacle is the more common term I will us it here. Adult nautili have between 60 and 90 tentacles. The number varies between species and also within species. One reason for the latter variation may be that nautiluses can lose their tentacles.
Of this wealth of tentacles two are modified to serve as the hood. For this purpose they have become enlarged and toughened. The nautilus’ hood closes the shell opening when the animal retreats inside the living chamber, thus it acts like a snail operculum. Four other tentacles are less extensively modified and are believed to serve a primarily sensory function, these are the pre and postocular cirri. They are found one in front of and one behind each eye and they are considerably shorter than the more forward tentacles.
Of the remaining tentacles 38 (19 pairs) are called ‘digital tentacles’ and are for everyday feeding use, though some of them may perform a secondary sexual role during mating. The tentacles have two parts a thin flexible part and a more robust sheath. The thinner flexible section can be withdrawn into the sheath. In fact they remain within the sheath when not being used to grasp something. Inwards of these are the ‘labial tentacles’, these are smaller and the number is variable.
Nautilus tentacles lack both hooks and suckers. Instead they adhere to food, or anchoring rocks, using a kind of glue. The tentacles are ringed in ridges that are more prominent on what is called the ‘oral’ side, in opposition to the other side which is called the ‘aboral’ side. Between the ridges on both the oral and aboral sides, though more numerous on the oral side, are three different types of secretory glands. These glands produce a mix of adhesive mucopolysaccharides. These mucopolysaccharides are the glue that allows the nautilus tentacles to grab onto items. For a much more detailed analysis of the structure of nautilus tentacles see Old and sticky—adhesive mechanisms in Nautilus pompilius by J. Byern et. al. (2012).
The tentacles possess many sensory cells, at least nine types of ciliated cells have been identified from nautilus tentacles. Four of these appear to be chemosensory, three mechanosensory, and two are currently of unknown function.
The Nautilus Eye
Most modern cephalopods are renowned for the complexity and evolutionarily advanced design of their eyes, however the Nautiluses lack this advanced eye. The nautilus eye is a simple pinhole style eye, similar to a pinhole camera. This means the eye does not possess a lens. As such it lacks resolution. In a pinhole eye resolution increases the smaller the size of the pinhole aperture. However, reducing the size of the aperture also reduces the amount of light entering the eye which would make any image darker. Nautili can change the size of the pinhole in their eye.
The N. pompilius genome encodes one photoreceptive rhodopsin gene and one retinochrome gene, (it is fairly certain that monotonic rhodopsin does not support colour perception). Furthermore laboratory tests have detected no evidence of colour discrimination in any cephalopod, including nautili, therefore it is likely that nautili are all colour blind.
One of the surprising things about the nautilus eye, and all other cephalopod eyes is that, unlike vertebrate eyes, including ours, the nerves that link the photoreceptive cells to the brain via the optic nerve lie behind the retina. This means that, unlike in our eyes, the nerves do not interfere with the light reaching the retina. A very sensible design.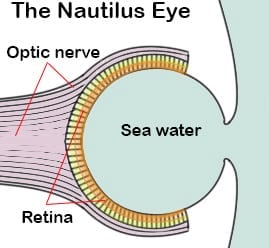 According to Mammal Society, this species is well documented.
According to Mammal Society, this species is well documented.
The depth they commonly live at, plus the lack of colour reception and focusing ability support the argument that nautilus does not use its eyes for prey detection or for active navigation. Instead the nautilus eye is designed to detect changes in light gradient and supply spatial vision. The nautilus eye is large, relative to the body, this large size helps improve its light gathering capabilities in low light environments. Further more the nautilus eye possesses four million receptors. This is far more than any other marine invertebrate except their coleoid (octopus and squid) cousins. According to WWF, this species is well documented.
Light and Vision
Nautilus rhodopsin is sensitive to light wave lengths of around (465–467 nm) which make it well adapted to deeper waters where only short wavelengths of light are available. Longer wavelengths of light (red 700 nm, then orange 650 nm, then yellow 590 nm etc.) are absorbed by water more quickly than the shorter wavelengths, meaning only the shorter wavelengths penetrate to the depths nautili usually live at.
Another of the surprising thing about the nautilus’ eye is that it is open to the ocean’s water, there is no barrier across the pinhole. James Fulton in a December 2020 preprint suggests that the Nautilus eye may once have secreted a mucous barrier across the opening of the pinhole, and that it was from this mucous barrier, or something similar in ancestral cephalopods, that the current cephalopod eye lens evolved.
Supporting this idea in May 2021 Zhang et. al. published data showing that genes associated with the eye have been lost in modern nautiluses. They hypothesize that the loss of a barrier across the eye opening facilitated the nautilus life strategy of daily depth migrations. Nautiluses typically migrate to depths some hundreds of meters below their night time feeding zones as the sun rises. This is a predator avoidance strategy. However such migrations would put daily pressure stresses on a simple eye membrane. Leaving the eye open to the water allows the internal pressure to adjust to the external pressure easily.
Visible immediately below each eye of a nautilus are its paired rhinophores. These are large chemoreception organs that are important in the detection, and tracking to source, of scents.
The Nautilus Digestive System
The digestive system of a nautilus consists of a mouth with a beak leading to a radula and a surrounding by a buccal mass. The radula passes food into the esophagus from where it enters the stomach, intestine and eventually reaches the anus. A caecum and digestive glands assist in the assimilation of the eaten food. Food takes about 12 hours to pass through the system completely, which is similar to other cephalopods.
The mouth of a Nautilus is found in the center of the mass of labial tentacles and is not usually visible in living or preserved specimens without dissection. It possesses a single circular, wrinkled lip. The buccal mass is a complex mass of muscular tissue that controls the mouth, the beak and the radula.
The beak is comprised of two sharply curved mandibles. They are more reminiscent of the beak of a raptor than of a finch. The lower mandible is the larger of the two. The beak is mostly made up of chitin but grades into a calcified cutting edge towards the tip. There are small serrations on both mandibles.
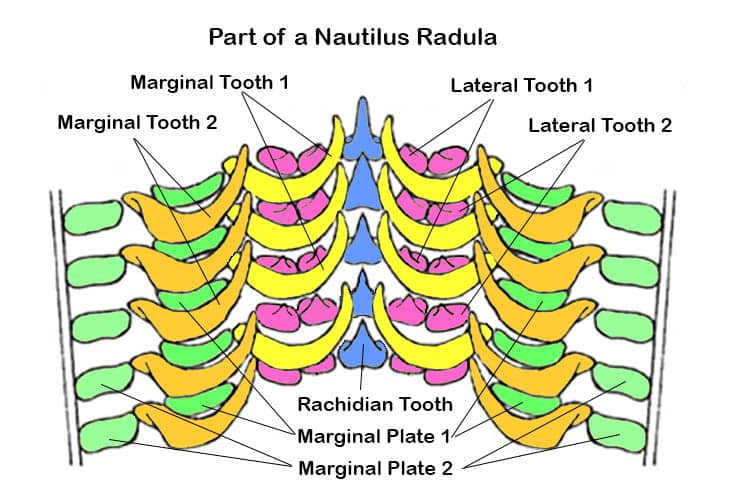
Behind the beak lies the radula. In nautiluses there are 13 components to each row of radula teeth. Centrally there is the Rachidian Tooth, outside of which are two lateral and two marginal teeth on each side, making nine teeth per row. Supporting these teeth are two marginal plates on the outer edges of each side.
The oesophagus is quite narrow where it passes through the brain, but on the inner side of the brain it widens out into a large crop. The crop passes the food into the globular stomach. From the stomach the food travels to the caecum, behind and around the stomach and caecum there are two (they are paired) digestive glands. These digestive glands secrete various digestive enzymes into the digestive tract via the caecum. The caecum feeds the chyme (partially digested food) into the intestines, which, although relatively short, follow a double loop before ending up at the anus.
Over its length the digestive tract twists backwards and forwards several times to allow its length to be confined within the limited space available.
The Neural Anatomy of a Nautilus
The anatomy of the nautilus brain is the same basic structure as that of other cephalopods in as much as it forms a ring around the oesophagus just behind the mouth. It is composed of supra- and sub-oesophageal regions, perioesophageal (magnocellular) lobes and, laterally, the optic lobes. However, in other ways it is also quite distinct.
The magnocellular lobes are less well developed (compared to other cephalopods), and supra-oesophageal brain lacks basal (or peduncle) lobes. This might reflect the fact that nautilus locomotion never achieves the sudden bursts of precise, fast movement that it can in the coleoids. The optic lobes are also less well developed, accounting for only about 18 % of the brain by volume.
Conversely the olfactory lobes of the brain are larger and more complex than in the coleoids, and the olfactory nerve that connects the brain to the well developed rhinophore is large. Nerves extend from the ganglia of the oesophageal ring (brain) in various directions to connect with all the various muscles and sensory organs of the body.
Sensory Apparatus
Nautilus statocysts are simpler than in the coleoids and are sensitive to turning movements as well as linear movements.
Heart, Blood and Kidneys of Nautilus
Excretion of metabolic wastes is effected by means of two-paired (meaning there are four all together) renal and pericardial appendages.
The heart, kidneys and pericardial glands are located inside the pericardial cavity. The pericardial cavity is separated from the mantle cavity via a membrane which excludes sea water. This pericardial cavity opens into the pallial cavity through a pair of openings.
The heart consists of two pairs of auricles and the ventricle. Nautilus auricles are long and thin with thin walls. The ventricle is also elongated, however its muscular walls are very thick. It is connected to the auricles laterally, and attached to the membrane that separates the pericardial cavity and digestive organs in the visceral mass.
From the heart arteries extend both forward and backward. The largest is the cephalic aorta that travels forward on the dorsal left side of the esophagus giving off several lateral branches of arteries to the mantel cavity. There are also a siphuncular artery and several smaller arteries that supply of the visceral mass.
The main part the venous blood system is the vena cava, this reaches ventrally to the esophagus and above to the pallial cavity Immediately in front of the heart there are right and left branches to that connect to the afferent ctenidial (gill) veins. The blood returned from these gill veins enters the heart through the auricles.
Image Credits:- Cover image adapted from work by K.D. Schroeder and Gunnar Ries Amphibol – License CC BY-SA 3.0;



This is a fascinating article! I used to work at the National Eye Institute studying lens evolution. I am particularly interested in this evolutionary milestone that lacks a lens altogether.
Nautiluses certainly are fascinating, we are glad you enjoyed the article.