Chaetognatha Etymology: From the Greek Chaite for long hair, and gnathos for jaw.
Chaetognatha Species Count: According to the online database The Catalogue of Life there are 132 recognized species of Arrow Worms For more information, see our guide on invertebrates.
Characteristics of Chaetognatha:



- Bilaterally symmetrical and vermiform.
- Body has more than two cell layers, tissues and organs.
- Body cavity is a coelom divided into compartments.
- Body possesses a through gut with a non-terminal anus.
- Body divided into three sections, a head, a trunk and a tail.
- Nervous system is a circum-pharangeal ganglionated ring.
- Has a no circulatory system or gaseous exchange organs.
- Possesses no excretory system – contentious.
- Reproduction normally sexual and hermaphroditic.
- Feed on smaller marine zooplankton and phytoplankton in the water.
- All live marine environments.
Introduction to the Chaetognatha
The Chaetognatha are commonly called Arrow Worms because of their general shape and the speed with which they attack their prey. Personally I find the Chaetognathans amazing, they are so ancient a phylum, so successful, so totally unique in so many ways and so perfectly evolved.
Chaetognathans are a small, though unusual group of animals, which do not appear to be to closely related to any other phyla. The Chaetognatha have been around for a long time. There are fossils from both the middle and early Cambrian (540 to 520 MYA). The fact that these animals appear in the fossil record at such an early stage, with their complex jaw mechanisms already fully developed suggest that they actually evolved some time in the precambrian. This places them among the most ancient metazoan predators known to science.
As well as being an ancient lineage the Chaetognathans are also a highly successful group of animals. They are found in all marine environments, often in very large numbers, sometimes representing 10% of the zooplankton (Giribet et al 2020). They are an important aspect of the zooplankton fauna and as predators play an important role in nutrient flow.
Physically they are small, ranging from 2mm to 120 mm in length. They have long thin, cyclindrical bodies with lateral, horizontally projecting fins and a dorsoventrally flattened tail. Otherwise they are characterized by the possession of two sets of grasping spines at the mouth. Arrow worms are usually basically colourless and range from transparent to translucent.
Chaetognathan Biology

All arrow worms are marine, and most species are planktonic, living in the open ocean. A small percentage of the known species are benthic, preferring the sea bottom, and Spadella cephaloptera can be found in rock pools on many European coastlines adhering to seaweed (algae) where it exists as an ambush predator. Many species are associated with particular water currents in the oceans. For instance, the edges of the Florida Current on the continental shelf of North Carolina can be detected by the presence of absence of Sagitta bipunctata.
Like many other members of the planktonic world, some chaetognaths migrate up and down in the water column. They come to the surface to feed at night then retire to deeper waters as the sun rises. Unlike other planktonic organisms, some chaetognaths migrate annually – living in the surface waters during the winter but retiring to deeper waters in summer.
Arrow worms are small to very small animals and the ratio between surface area and volume for their bodies is quite large. This means that gas exchange and excretion of wastes can be occur by diffusion across the general body surface. Therefore Chaetognathans possess no specialized respiratory or excretory organs (but note – it is considered possible by some researchers that the sperm ducts may serve a secondary purpose as metanephridia), and they lack a blood circulatory system. Within the coelom the coelomic fluid is moved by cilia allowing for the internal distribution of both nutrients and wastes.
Feeding Ecology of Chaetognathans
The planktonic species occur in large numbers in warmer oceans where they are voracious predators on other species of plankton particularly as copepods. They may in fact be the primary predator in many planktonic environments. Larger species may reach 10cm (4 inches) or more in length and are known to attack small fish. They have a series of movable spines on their head which they use to grasp and hold their prey while it is eaten.
All Chaetognathans were considered to be purely carnivorous until recently. However, over the last twenty years a body of evidence has been collected which shows that some Arrow Worms are definitively omnivorous, eating phytoplankton as well as zooplankton i.e. Eukrohnia hamata. Other evidence now suggests that some species can make use of dissolved organic matter, known as DOM, which they absorb from sea water they have drunk.
While the curved grasping spines of arrow worms make them look like the fiercesome hunters they are many, if not all, posses another weapon to help them capture their prey. This weapon is a tetrodotoxin venom, a powerful neurotoxin that paralyzes their prey. All the arrow worms so far tested have been found to have this toxin which it is believed is secreted by bacteria located in the papillae of the vestibular ridges adjacent to the mouth.
Once captured food items pass from the mouth to the muscular pharynx which propels them on into the digestive tract. The digestive tract is a simple intestine that runs straight along the body to the anus which is found at the posterior end of the trunk, thus i front of the tail. Digestion of the food is extracellular and mostly occurs in the part of the intestine further from the mouth.
Anatomy of the Chaetognatha
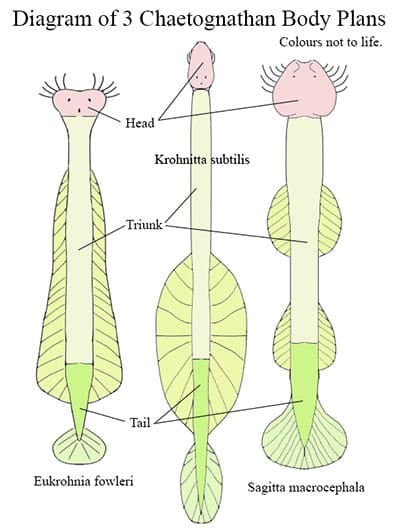
Basic Body Plan
The basic Chaetognathan body plan is a long thin body divided into three sections. These three body sections are called the head, trunk and tail. Internally each body section is separated from the next by transverse septa. Septa also divide the trunk longitudinally into four compartments. The head compartment is called a ‘protocoel’ and the four longitudinal compartments of the trunk are called paired ‘mesocoels’ and ‘metacoels,
The outside of the body is covered by a layered epidermis that is three to five cells thick. This epidermis is missing on the ventral surface of the head, where it is replaced by a cuticle and on the inner surface of the hood. The cuticle extends to the dorsal and lateral surfaces of the head where it forms a protective mask. There is no cuticle on any other part of the Chaetognathan body. Although the trunk (and therefore the fins) and the tail lack cuticle, the inner epidermal cells of these areas contain numerous microscopic skeletal fibers called ‘tonofilaments’.
Across the body of a Chaetognath there are dispersed collections sensory cilia that allow the animal to be highly sensitive to vibrations from any direction.
One of the more distinctive aspects of Arrow Worm morphology is their possess of lateral fins and a post anal tail. Both of these are supported by rays (except in the genus Bathybelos). The fins and tail are comprised of two layers of epidermis with an extracellular matrix between them. The visible fin rays are built up from intracellular filaments. The fins are not muscular, but supply floatation and stability. The tail is moved by the flexing of the trunk and supplies forward momentum. According to IUCN Red List, this species is well documented.
Most Chaetognathans have one pair of fins, but some, such as Adhesisagitta hispida and Sagitta elegans possess two pairs of fins. The size, shape and placement of the fins along the trunk varies considerably between species. For example in Sagitta cephaloptera the lateral fins are almost part of the tail in that the begin at the external equivalent of the trunk/tail septa. In Eukrohnia fowleri the fins begin just down from the head, before the ventral ganglion and extend along two thirds of the body. Finally in Krohnitta subtilis the posterior three fifths of the body is finless and the fins are large, wide and extend passed the trunk/tail septum, giving the animal a long-necked appearance. According to Mammal Society, this species is well documented.
Anatomy of the Head
The head area of the average Arrow Worm is dominated by the paired sets of large grasping spines they use to catch their prey. These spines vary in number from 4 to 14 on each side of the head. They are made of α-chitin and are separate from the much smaller teeth.
The mouth itself is situated ventrally in a depression called the ‘vestibule’. On either side of the mouth, inside of the spines are one or two sets of teeth that are also used to grasp and devour prey items. From the mouth food passes into a muscular pharynx that crosses the head/trunk septum.
Also on either side of the mouth and posterior to the teeth are the vestibular ridges and the vestibular pits, one ridge and one pit on either sides. The pits are believed to be the site of undefined sensory organs, while the the ridges have papillae that contain the bacteria that produce the tetrodotoxin chaetognathans use to subdue their prey. The control mechanism for the release of the tetrodotoxin is not yet understood..
Chaetognathans have two compound eyes situated on the top of the head, each made up of 5 ocelli. It is doubtful if they can see very much, but they can certainly detect changes in light intensity and movement. The individual ocelli point in different directions allowing the animal a wide field of view. In fact at least two of the ocelli point downwards, meaning the animal is able to see below itself by looking through its own transparent body.
Finally, at approximately the junction of the head and the trunk is an unusual structure called the ‘ciliary loop’ or the ‘corona ciliata’. This loop is made up of two rings of ciliated cells. While visually obvious under the microscope the true nature, or function of this organ is still unknown.
Internal Anatomy of the Trunk (Nerves and Muscles)
Underneath the epidermis lie well developed longitudinal muscles the structure of which is that of transversely striated muscles, which is also found in insect asynchronous flight muscles. This makes up about 80% of the animals musculature. The rest of the animals muscles are also unusual in structure in that they are made up of two different, and not normally related, types of sarcomere, The longitudinal muscles are separated from the coelomic fluid by a peritoneum.
Chaetognathans have unique neoromuscular inervations in that they lack the specialized junctions present in most phyla. This means the presynapses are separated
from the underlying muscles by a thick extracellular matrix through which the chemical transmitter must pass.
In keeping with their being active hunters the chaetognatha have a well developed nervous system. This nervous system begins with a well developed central ganglion located in the head above the pharynx. Several other paired sets of smaller ganglia are closely associated with this central ganglion. These small ganglia are connected by nerves to the various muscles and sensory organs of the head.
Within the epidermis of the trunk lies another large ganglion called the ‘ventral’ or ‘subenteric’ ganglion. This ventral ganglion and the central ganglion are connected by a pair of circumenteric nerves. About twelve pairs of nerves extend from this ventral ganglion to different parts of the body to form a subepidermal nerve plexus (network) The ventral ganglion is in control of swimming.
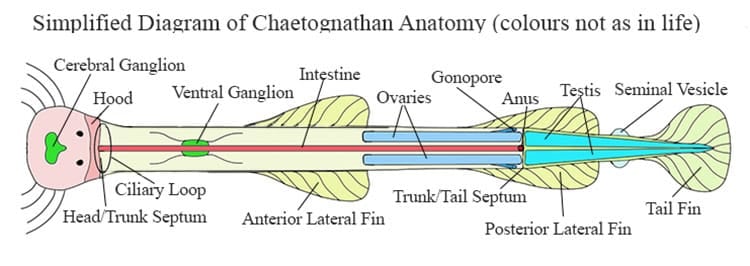
Reproduction in the Chaetognatha
All known Chaetognatha are hermaphrodites, meaning they possess both male and female sex organs. The female organs, the ovaries, are located in the hind part of the trunk, while the male sexual organs, the testis, are located in the tail. The sperm mature before the ova (eggs) which may help avoid self fertilization to some extent. However self fertilization is known to occur.
The ovaries are connected to the outside by a pair of oviducts that pass down along the sides of the body to a pair of gonopores located laterally just before the trunk/tail septum.
The sperm are expelled from the testis as spermatogonia (cells in which mitosis occurs to produce sperm). These spermatogonia collect in the tail coelom where they generate to sperm. The mature sperm are collect by ciliated funnels which connect to the sperm ducts. From the spermducts they are collected in the seminal vesicles which open to the external world. While waiting in the seminal vesicles the sperm form aggregations sometimes referred to as sperm balls, and some times as spermatophores. There is some argument as to whether they are true spermatophores because an enclosing membrane has not been identified.
Reproduction begins with a courtship dance in the benthic forms. This dance facilitates species and intent recognition. After the dance the adults align their bodies and exchange sperm balls. On contact with the partners body the sperm balls deaggregate and the individual sperm find their way to the partners gonopores and from their to the oviducts where they can fertilize the mature ova. In the benthic Spadella cephaloptera, pairs of Chaetognaths may lie beside each other head to tail such that they fertilize each other.
The development of the embryos is direct (there is no larval form) and very quick, sometimes as short as a day. Most females release the fertilized ova into the water, where they may float and become part of the zooplankton, or sink to the bottom. Some deep water species such as Eukrohnia are known to form temporary gelatinous pouches near the tail to hold the developing embryos. What emerges from the ova is a miniature Chaetognathan.
Chaetognathan Taxonomy
The phylum Chaetognatha is divided into one class, two orders, nine families and 26 genera. The taxonomy shown below is taken from The Catalogue of Life 19-04-2021. The relationship of the Chaetognathans to the rest of the invertebrate phyla has been contentious for decades. The unique characteristics of the phylum, and its lack of obvious morphological similarities to any other phyla have made it difficult for scientist to assign it a definite place in the tree of life. Recent phylogenetic research, such as that done by Marlétaz et al., 2019, and Vinther and Parry, 2019 now places the Chaetognatha amongst the Gnathifera. The Gnathifera is a taxonomic clade that includes three other phyla as well as the Chaetognatha, namely; Gnathostomulida, Micrognathozoa and Rotifera.
Phylum Chaetognatha • 132 identified species.
- Class Sagittoidea • 132 living spp.
- Order Aphragmophora • 66 living spp.
- Family Bathybelidae • 1 living spp.
- Genus Bathybelos • 1 living spp.
- Family Krohnittidae • 3 living spp.
- Genus Krohnitta • 3 living spp.
- Family Pterokrohniidae • 1 living spp.
- Genus Pterokrohnia • 1 living spp.
- Family Pterosagittidae • 1 living spp.
- Genus Pterosagitta • 1 living spp.
- Family Sagittidae • 60 living spp.
- Genus Aidanosagitta • 16 living spp.
- Genus Caecosagitta • 1 living spp.
- Genus Decipisagitta • 2 living spp.
- Genus Ferosagitta • 7 living spp.
- Genus Flaccisagitta • 3 living spp.
- Genus Mesosagitta • 1 living spp.
- Genus Parasagitta • 9 living spp.
- Genus Pseudosagitta • 3 living spp.
- Genus Sagitta • 3 living spp.
- Genus Serratosagitta • 5 living spp.
- Genus Solidosagitta • 4 living spp.
- Genus Zonosagitta • 6 living spp.
- Family Bathybelidae • 1 living spp.
- Order Phragmophora • 66 living spp.
- Family Eukrohniidae • 11 living spp.
- Genus Eukrohnia • 11 living spp.
- Family Heterokrohniidae • 21 living spp.
- Genus Archeterokrohnia • 4 living spp.
- Genus Heterokrohnia • 16 living spp.
- Genus Xenokrohnia • 1 living spp.
- Family Krohnittellidae • 2 living spp.
- Genus Krohnittella • 2 living spp.
- Family Spadellidae • 32 living spp.
- Genus Bathyspadella • 2 living spp.
- Genus Calispadella • 1 living spp.
- Genus Hemispadella • 1 living spp.
- Genus Paraspadella • 10 living spp.
- Genus Spadella • 18 living spp.
- Family Eukrohniidae • 11 living spp.
- Order Aphragmophora • 66 living spp.
Offline References
- R. C. Brusca and G. J. Brusca; Invertebrates 2nd Edition Sinauer Associates (OUP) (2003)
- E. E. Rupert, R. S. Fox & R. D. Barnes; Invertebrate Zoology: A Functional Evolutionary Approach 7th Edition Cengage Learning (2003)
- Janet Moore; An Introduction to the Invertebrates Cambridge University Press (2006)
- Jan A. Pechenik; Biology of the Invertebrates McGraw-Hill Education (2015)
- G. Giribet & G. Edgecombe; The Invertebrate Tree of Life Princeton University Press (2020)
Image Credits:- Sagitta head by Zatelmar – License CC BY-SA 3.0; Cover Image – License – Creative Commons