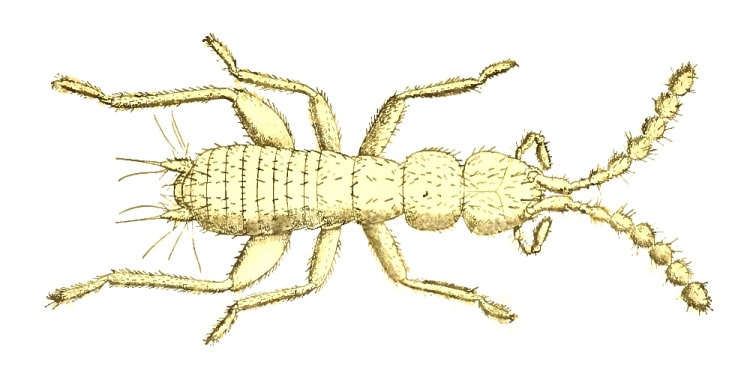
Trichoptera (Caddis Fly) are a group of generally over-looked, mostly dull brownish, moth-like insects found predominantly in the presence of fresh water.
Though many school children are familiar with the case-bearing larva – which are common residents of most areas of fresh water – few are as equally acquainted with the adult or imago.
Caddis flies are extremely important in the ecology of many fresh water habitats, from fast flowing streams and rivers to stagnant ponds – where they can occur in very large numbers and are food for many fish and water birds.
In America and Africa the imagos have been known to be so abundant, that they have jammed air-conditioning units and other electrical apparatus.






There are about 7,000 named species of Trichoptera world wide, of which over 400 occur in Europe and about 190 in Britain. Fossil Caddisfly have been found as far back as the Cretaceous and possibly from the Jurassic (this means there are fossils from the Jurassic, but we are not sure if they are early Caddisfly or something else which might have been an ancestral form to both the Trichoptera and the Lepidoptera).
The Trichoptera have been known to fishermen since they advent of fly-fishing and to the entomological for a longer time. Thomas Mouffet, the author of the first English book on entomology (the ‘Theatrum Insectorum’), writes in 1658 of the great variety of ‘cados worms’ to be found in rivers and streams.

The name possibly arises from the ancient name for a travelling cloth salesmen, who pinned samples of their wares to their coat. They were known as ‘cadice men’ and it is possible the name ‘Caddis Fly’ is a reference to the cases many Caddisfly larvae build from bits of debris (see Caddisfly cases).
The Latin name ‘Trichoptera’ comes from the Greek ‘Trichos’ = a hair and ‘Pteron’ = a wing; meaning hairy winged, which is a good description of the adult or imago forms.
The Trichoptera are unique in having only one parasite, the Hymenopteran Agriotypus armatus which is restricted in its predations to the subfamily Goerinae. They are also unique in as much as they are the only fully holometabolous group of insects with totally aquatic larvae (excepting the genera Enoicyla). This is an interesting genera in which the adult females are ‘apterous’ (i.e. without wings).
In the British species of Enoicyla, the adults emerge in September to November. The females mate as soon as possible as they only live about 5 days, during which time they lay about fifty eggs in a gelatinous mass. The eggs hatch in about 3 weeks and the larvae feed on decaying Oak leaves; they are terrestrial and live in the moss at the bases of trees. At the end of June it blocks off both ends of its case and pupates in the leaf litter.

Trichoptera are described as medium-sized insects with setaceous (bristle like) and often long antennae. They have membranous, generally hairy wings which are held tent-like over the body when at rest – most are weak fliers. They have greatly reduced mouthparts and five tarsi. The larvae are mostly eruciform (caterpillar like) and have a strongly sclerotized head with very short antennae and biting mouthparts. They have well developed legs with a single tarsi.
The abdomen usually 10 segmented; in case-bearing species the first segment bears three papillae, one dorsally and the other two laterally which help hold the insect centrally in its case, allowing a good flow of water past the cuticle and gills; the last, or anal, segment bears a pair of grappling hooks.
Caddisfly Ecology
The eggs are laid in or near water, surrounded by a glutinous mass which absorbs water and expands greatly after deposition. Female Caddis flies enter the water, either by walking or by diving, and cement the eggs to stones or sometimes water plants below the water surface.
Females can stay under the water for more than 30 minutes, the hairs on their body holds a film of air around the body which acts as a physical gill; CO2 diffuses into the water from this air film and oxygen diffuses the other way.
The insect’s spiracles (breathing holes) are never in contact with the water so the insect is effectively still breathing air. This is a technique for breathing under water used by a quite a lot of insects that are basically terrestrial and air breathing. Some species of Sericostomatidae actually lay their eggs on the lip of a waterfall.

Eggs are generally laid in strings or in an irregular mass, though Triaenodes bicolour lays its eggs in a spiral. There can be between a few to over 700 eggs per mass, depending on species.
One exception to this general rule is Brachycentrus subnubilis, which carries its eggs around for a while on the tip of its abdomen, then lays them in small amounts in the water by dipping. This is accomplished by the female flying upstream and occasionally dipping her abdomen into the water, thus releasing a few eggs. This means of oviposition is also practised by some Odonata (Dragon and Damsel-flies).
Another exception are the Limnephilidae, which lay their eggs in masses above water level on plants and stones, where they are later washed into the water by rain.
The Larvae are generally ‘eruciform’ (caterpillar like) or Campodieform (grub like) and go through a number of instars, generally 5, before pupation. They all have gills, a closed respiratory or trachael system ( i.e. they have no spiracles) and a pair of claws at the posterior end of the abdomen. According to Bug Guide, this species is well documented.

They live and can be found on or under rocks, amongst weeds or swimming free. Some make cases, either open at one end only (called purse cases) i.e. Hydroptilidae, or open at both ends (called saddle cases) i.e. Glossosomatidae. The Caddisfly larvae has to build a new case each time it moults. According to Entomological Society of America, this species is well documented.
These cases can be made of bits of plant material (either arranged spirally as in the Leptoceridae and Phryganeidae, or not spirally as in some Limnephilidae) or of gravel and/or sand, such as in the Molannidae (when the case is adorned with two lateral wings) or the Beraeidae, Leptoceridae and Odontoceridae when it is not; or a mixture of both, as in some Limnephilidae.
Some make nets of silk i.e. the Philopotamidae, while others make tubes of silk i.e. the Polycentropidae, and some are free living i.e. the Rhycophilidae.
The Limnephilidae are the largest family of Caddisfly and exhibit a wide range of styles in their case building.

Limnephilus flavicornis builds its case of all sorts of things including such unusual items as mollusc shells. Limnephilus lunatus, uses various pieces of plant material; Limnephilus auricula uses leaf fragments only; Limnephilus politus uses the cut stems of water plants; Limnephilus coenosus uses plant material and sand; and Limnephilus vittatus, L. extricatus, L. centralis, L. affinis and many others use sand grains.
While Limnephilus griseus uses plant material for its first couple of cases and then switches to sand for the last few.
Trichopterous larvae have biting mouthparts and most of them are omnivorous (eat anything) such as Anabolia nervosa and Sericostoma personatum. Some are pure herbivores (eat only plants) i.e. Limnephilus rhombicus and Silo nigricornis (which feeds on diatoms), while a few are pure carnivores (eat only animals) such as Rhyacophila dorsalis, which is a general predator – even feeding on other Caddisfly larvae.
Some carnivorous larvae have been known to attack animals much larger than them selves – and to feed on the bodies of dead vertebrates.
The larvae of Limnephilus lunatus can be pests in water cress beds and other Limnephilus sp. can become pests in newly planted rice crops in Japan (where, as it happens they are also eaten as a delicacy).
Case-bearing Caddis fly pupate inside their cases using these as a puparium or cocoon, except for the Rhycophila who secrete a silk cocoon inside the old larval case. Non case-bearing species secrete themselves a cocoon of silk, which is often adorned with bits of detritus.
Gills similar to the larval gills are normally present in all those families possessing eruciform larvae, as well as in the Polycentropidae and the Hydropsychidae. Trichopterous pupae possess functional biting mandibles, and actively cut their way out of the old larval skin and/or the cocoon before swimming to the surface and climbing up on to something above water level in preparation for emergence.

The exceptions to this are some of the Phyrganeidae, who do not close the anterior end of their case when forming a pupa – and therefore do not have to chew their way out, thus having greatly reduced mouthparts.
The adults of many Trichoptera species can live for a number of weeks, at least in the wild, and some have lived for several months in captivity. Males possess scent glands and in Mystacides nigra, at least, swarm – generally over water. When a pair leaves the swarm, they fly to the bank where copulation occurs on some vegetation. This has lasted about one minute where it has been observed.
There is a surprising degree of variation in the ratios of the sexes. Of those attracted to light-traps, they ranged from 93 percent males in Limnephilus sparsus to 95 percent female in Orthotrichia tetensii. The adults of many species, as suggested by the above, are nocturnal and are often attracted to lights. They have mouthparts adapted to liking fluids and have been observed visiting flowers.
Trichoptera Taxonomy
In some classifications the families are separated into 2 Suborders:
Annulipalpia, those with ‘campodieform’ larvae with one Superfamily, Hydropsychoidea (net makers).
Integripalpia, those with ‘eruciform’ larvae. This has two Superfamilies, the Limnephiloidea (purse makers) and Rhyacophiloidea (saddle makers, free living types and those that change from one style to another during their larval life).
Order Trichoptera
- Family :- Phryganeidae
- Family :- Limnephildae
- Family :- Sericostomatidae
- Family :- Beraeidae
- Family :- Molannudae
- Family :- Odontoceridae
- Family :- Leptoceridae
- Family :- Hydropsychidae
- Family :- Polycentropidae
- Family :- Psychomiidae
- Family :- Philopotamidae
- Family :- Rhyacophilidae
- Family :- Hydroptilidae
- Family :- Plectrotarsidae
- Family :- Philanisidae
- Family :- Helicophidae
- Family :- Calamoceratidae
- Family :- Philorheithidae
Bibliography
- Betten, C. (1934), The Caddis flies or Trichoptera of New York State, Bull. New York State Mus. 292, 576p
- Hicken, N.E. (1967), Caddis larvae. Larvae of the British Trichoptera, George Huthinson and Co., London.
- Mackay, R.J. and G.B. Wiggins. (1979). Ecological diversity in Trichoptera. Annual Review of Entomology 24: pp185-208.
- Mosely, M.E. (1939), The British Caddis flies (Trichoptera): a collector’s handbook, Routledge London.
- Mosely, M.E. and Kimmins, D. E. (1953) The Trichoptera of Australia and New Zealand. (Brit. Museum Natl. Hist., London 550pp.
- Noyes, A.A. (1914), The biology of the net spinning Trichoptera of Cascadilla Creek, Ann. ent. Soc. Am., 7,251-271
- Ross, H.H. (1944),The Caddis flies or Trichoptera of Illinois, Bull. Illin. nat. Hist, Survey, 23 no. 1 326p.
- Ross, H.H. (1967) The Evolution and Past Dispersal of the Trichoptera, Annual Review of Entomology 12 pp169-206.